|
|
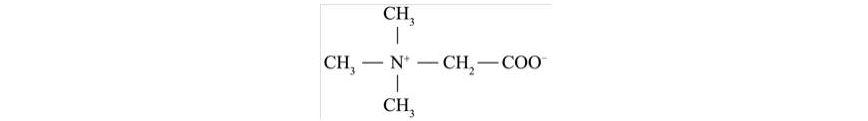
甜菜堿概述發表時間:2019-06-21 17:13 1 甜菜堿簡介 甜菜堿的發現已有50多年的歷史,最早從甜菜制糖產生的廢蜜中分離而得,又稱三甲基甘氨酸等,是一種氨基酸衍生物,電荷在分子內分布呈中性,三個甲基活性無差異,因此它是高效的甲基供體[1];甜菜堿是一種季銨型生物堿,具有親水和親脂性,20℃水中溶解度為160g/100g,親脂性使其在生物體中能夠容易透過磷脂分子層,是公認的在細胞中起著無毒滲透保護作用的細胞相溶性物質。甜菜堿具有雙電荷(兩性)結構特征,在堿性條件下呈環狀結構,酸性條件下呈開環狀,并具有較好的穩定性和抗氧化能力,化學結構與氨基酸、膽堿相似,具有多種生物學功能。廣泛應用于飼料工業、化工、印染、日化、醫藥、食品等多種領域。甜菜堿結構式如圖1所示。
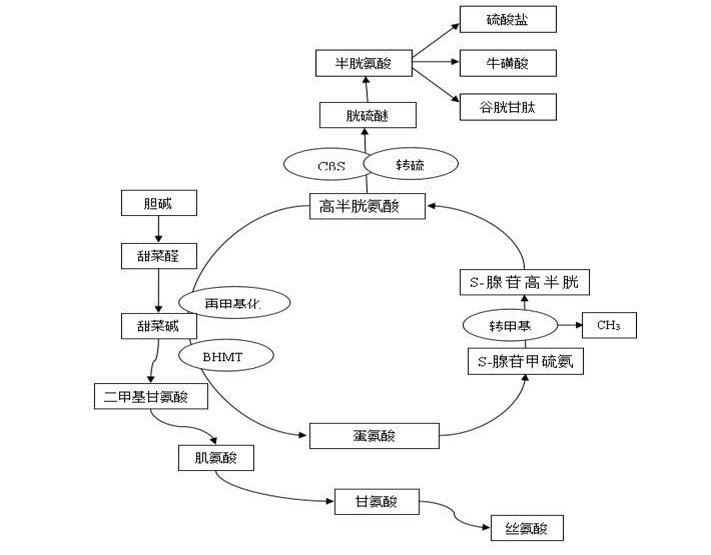
圖1 甜菜堿結構式 2 甜菜堿的吸收與利用 2.1 甜菜堿的吸收 甜菜堿吸收與氨基酸類似,主要通過甜菜堿-γ-氨基丁酸轉運系統和氨基酸轉運系統A進行轉運[2-5]。外源甜菜堿主要在動物十二指腸和空腸吸收,十二指腸的吸收效率高于空腸,后依靠小腸上皮細胞的Na+和Cl-泵進行主動轉運、不依賴Na+泵途徑及被動擴散進行轉運[6, 7]。 2.2 甜菜堿的利用 2.2.1 甜菜堿的代謝 甜菜堿在動物體內的利用率很高,健康動物即使攝入高劑量甜菜堿,也較少會通過排泄途徑排出,幾乎可全部被機體利用[8]。 在動物肝、腎、肌肉、眼睛等組織中均存在與甜菜堿代謝相關的酶,其中肝臟是主要代謝地[9]。甜菜堿在肝臟中受甜菜堿-高半胱氨酸-甲基轉移酶(Betaine homocysteine methyltransferase,BHMT)及胱硫醚-β-合成酶(Cystathionine-β-synthase,CβS)調控[10],且這些反應可通過蛋氨酸循環(如圖2)參與機體甲基代謝。 HMT作為甜菜堿轉甲基酶,是一種含鋅金屬酶,可催化甜菜堿為高半胱氨酸提供一個甲基以便再合成蛋氨酸,該過程稱為蛋氨酸的再甲基化[11],也是蛋氨酸再甲基化的重要途徑之一,甜菜堿則代謝為二甲基甘氨酸,再進一步代謝成肌氨酸、甘氨酸、絲氨酸等次級代謝產物。這對機體提高蛋氨酸利用率、解除高半胱氨酸的毒性和生成S-腺苷甲硫氨酸(S-adenosylmethionine,SAM)都具有重要意義[9]。有研究指出,日糧中添加甜菜堿可顯著提高肝臟中BHMT的水平[12],加速蛋氨酸的再甲基化,提高蛋氨酸及SAM含量[13];且能提高CβS水平,加速轉硫途徑[14],促進胱氨酸、牛磺酸、谷胱甘肽等生成。甜菜堿對蛋氨酸再甲基化及轉硫途徑的影響與添加劑量相關:低劑量甜菜堿的主效應為促進蛋氨酸的再甲基化;而高劑量添加甜菜堿可加速蛋氨酸的再甲基化及轉硫途徑[12, 14]。
圖2 甜菜堿代謝的轉甲基作用 2.2.2 在組織中沉積 若甜菜堿不經上述途徑代謝,則在肝、腎及肌肉等組織中沉積。汪以真等[15]研究指出,日糧中不額外添加甜菜堿,在豬的肝、腎及肌肉組織均能夠檢測到甜菜堿。Peuranen等[16]指出,日糧分別添加0、250、500、1000mg/kg甜菜堿,豬肝中甜菜堿含量隨著甜菜堿添加量增加而呈線性遞增。甜菜堿在各組織中沉積效率不同,主要沉積于肝臟和腎臟,其次是肌肉[17]。 3 甜菜堿的生理功能 3.1 調節滲透壓 滲透壓平衡對動物體蛋白質表達;氨基酸及氨類、碳水化合物及脂肪酸代謝;原生質膜運輸和基因表達等具有重要意義[18]。當機體細胞環境滲透壓發生劇變,細胞主要通過積累低分子量無機離子(Na+、K+等)和有機滲透壓調節物(甲基化堿基、氨基酸及其衍生物、糖醇等)來調節滲透壓;而無機離子對滲透壓調節能力有限,且高濃度的無機離子容易破壞機體蛋白質結構、降低酶活性、影響細胞正常代謝[19]。甜菜堿作為雙極性季胺鹽,具有類似電解質的特征及高溶解度,可沉積在細胞及細胞器中,在滲透壓激變時發揮對細胞的滲透壓保護作用。甜菜堿不僅不會干擾細胞正常代謝,且能夠穩定細胞正常體積,提高細胞中自由水的含量,緩解高滲應激對細胞增殖的影響[20],另外,甜菜堿能夠穩定細胞中蛋白及其他物質的結構和功能[21, 22]。 外源甜菜堿攝入能在腎臟中積累[23],保護細胞、減少高濃度電解質和尿素對細胞結構和功能的損傷[24, 25]。體外研究表明,甜菜堿可緩解由滲透壓劇變引起的肝臟巨噬細胞腫瘤壞死因子α釋放、前列腺素形成及環氧化酶-2的表達等造成的細胞損傷,穩定免疫功能[26]。甜菜堿能緩解由高滲環境引起豬肺動脈上皮細胞的程序性死亡,增加細胞對滲透壓變化的適應,減少在滲透激變時對細胞和器官的損傷[27]。Kettune等[28]研究發現,在飲水中添加0.2%甜菜堿能提高肉雞小腸上皮細胞的滯水能力,改善小腸上皮細胞結構,緩解高滲應激對肉雞的影響。 3.2 供甲基 甲基(-CH3)是合成蛋氨酸、肉堿、肌酸、磷脂、腎上腺髓質、核糖核酸、脫氧核糖核酸等物質的必須基團。機體自身不能合成甲基,需外源提供[29]。甜菜堿、膽堿和蛋氨酸是動物體主要的有效甲基供體。甜菜堿含有3個極性甲基,是有效的甲基供體,且能通過BHMT的催化作用增加蛋氨酸循環次數,增強機體甲基代謝,加速DNA等的加工和修飾等過程[30]。 3.3 調節蛋白代謝 甜菜堿能使動物肝臟和肌肉中的粗蛋白、核糖核酸、RNA與DNA的比值顯著提高,血清尿酸明顯下降。甜菜堿參與合成肌酸、磷脂、腎上腺素、核糖核酸、脫氧核糖核酸等多種重要生理功能物質,促進機體蛋白質的合成并降低蛋白質的降解,促使組織中蛋白質的沉積[31-34]。蛋氨酸不僅可合成機體蛋白質,同時還在蛋白質代謝中起重要作用[35]。甜菜堿可節約供甲基部分的蛋氨酸,降低蛋氨酸的供甲基消耗量,提高其利用效率[34,36-37]。 3.4 調節脂肪代謝 豬[38]、鴨[39]、蛋雞[40]及肉雞[41, 42]的研究表明,日糧中添加甜菜堿能有效的降低體脂沉積。甜菜堿可抑制脂肪酸合成相關酶及mRNA表達,從而減少脂肪沉積[43, 44];Xing等[44]研究指出,甜菜堿降低脂肪合成酶相關基因表達與基因的甲基化有關,提示甜菜堿可能通過調控基因甲基化而影響脂肪代謝合成酶基因表達。在降血脂與保肝作用的方面研究表明:甜菜堿顯著提高動物血清及肝中磷脂及極低密度脂蛋白的含量,促進動物肝中脂肪的遷移、降低肝臟中甘油三酯含量的作用。 甜菜堿可為肉堿合成提供甲基,促使肝臟中肉堿的合成量增多[34],使脂肪酸的運載增強,促進細胞線粒體內脂肪酸的β-氧化,并由此反饋性地增強了不同階段脂肪分解酶的活力,加強了脂肪的分解,使體內脂肪沉積相對減少。同時,甜菜堿可調控機體的激素水平和生長因子等會從而調節機體脂肪的分解和合成代謝[38]。 3.5 抗氧化 甜菜堿可能經以下途徑參與機體抗氧化:(1)參與蛋氨酸循環,提高蛋氨酸及SAM水平,促進胱氨酸、谷胱甘肽等物質生成,而這些代謝產物在機體抗氧化過程中發揮重要作用,尤其是GSH可直接清除體內自由基等,對機體抗氧化及細胞膜的穩定意義重大[46-47]。(2)調節機體滲透壓,對細胞、酶等結構和功能起穩定、保護作用。(3)甜菜堿能夠快速通過磷脂雙分子層且能與其產生特定生物學交互作用,減少細胞膜脂質過氧化[48]。 Kim等[49]研究指出,甜菜堿能顯著提高小鼠肝中谷胱甘肽含量和還原型與氧化型谷胱甘肽的比值。Alirezaei等[50]和Kheradmand等[51]報道,甜菜堿可提高小鼠肝臟GPX活性,降低肝臟丙二醛含量。Wang等[52]研究發現,甜菜堿提高了大鼠肝臟中谷胱甘肽過氧化物酶活性。另有研究指出,甜菜堿可提高肉雞胸肌中SOD、GPX等活力,緩解飼喂低蛋氨酸日糧引起的氧化應激,減少脂質過氧化產物生成[53]。 3.6 抗應激 甜菜堿可調節機體滲透壓、平衡水分代謝、保護腸道結構、增強免疫力、提高抗氧化功能及抑制中樞神經系統對應激的敏感性等,提高動物對應激環境的適應性和抵抗力。 高半胱氨酸是一種興奮性氨基酸,也是腦內γ-氨基丁酸(GABA)合成酶-谷氨酸脫羧酶(GAD)的抑制劑,而GABA是中樞神經系統中很重要的抑制性神經遞質。甜菜堿促進體內高半胱氨酸向蛋氨酸的轉化,降低體內高半胱氨酸的含量,減輕高半胱氨酸對GAD的抑制,有利于腦內GABA的合成,使中樞抑制作用加強,降低中樞對應激的敏感性,進而起到鎮靜、緩解應激的作用。 熱應激是關注及研究較多的一種應激,易導致動物精神不振、體溫升高、熱性喘息、呼吸加快、酸堿平衡紊亂、水分過度攝入及代謝紊亂、腸道結構損傷、免疫機能下降、采食量降低、生產性能降低、甚至死亡等[54, 55]。球蟲感染普遍存在于肉雞養殖場,導致肉雞發病和死亡、損傷腸道,阻礙肉雞生長發育,降低生產性能[56, 57]。 日糧中添加甜菜堿能減少熱應激肉雞的水分消耗,提高生產性能,緩解熱應激引起的過度飲水而導致飼料消化率下降[58];提高熱應激肉雞免疫功能,降低死淘率,減少無機離子的積累[59];改善熱應激肉雞酸堿平衡及水分在體內的滯留能力[60]。 Kettunen等[7]報道,甜菜堿降低了球蟲感染肉雞腸道的隱窩深度與絨毛長度比值,降低腸道損傷,提高腸道吸收表面積,增加飼料利用率。甜菜堿可改善球蟲感染肉雞生產性能[61],提高肉雞的飲水量及水在肉雞體內滯留能力,但甜菜堿的抗球蟲作用與球蟲的種類有關[62]。 參考文獻(略) |